根据中国海洋局公布的《2016 年中国海洋经济统计公报》,2016 年全国海洋生产总值达到7 万多亿元,占国内生产总值的9.5%,航运业以及海洋相关产业已经成为当今经济发展的重要支柱。海洋腐蚀是海洋工程面临的重大难题,世界腐蚀组织(WorldCorrosion Organization,WCO)估计每年海洋腐蚀损失约为0.8 万亿美元,占到全球腐蚀总量(2.4 万亿美元)的1/3,高于所有自然灾害损失的总和。
海洋腐蚀在造成大量经济损失的同时,还会引起严重的人员安全与环境污染问题,英国曾报道“亚历山大基定号”海洋平台桩腿焊缝因海水腐蚀形成裂纹,在海浪的反复作用下裂纹不断扩大,最终平台倾倒并造成123 人遇难。美国海域发生过的最严重的油污染就是因为“深水地平线”钻井平台海底阀门受海水腐蚀而失效爆炸,导致3 个月内约400 万桶原油泄漏入海。我国海洋腐蚀研究起步较晚,目前多数海洋工程结构物与载体处于欠防护状态,腐蚀损失巨大且安全堪忧,海洋腐蚀已成为我国海洋经济大发展中必须攻克的关键问题。
海洋环境下金属材料的腐蚀包括电化学腐蚀和污损生物腐蚀,每年海洋金属材料因腐蚀而造成的损失中,约有20%是由微生物腐蚀引起的。微生物在金属表面的腐蚀还会与磨损、气蚀、冲蚀等因素协同,导致材料表面快速剥落,使包括船舶与海洋平台在内的海洋设施的可靠性面临严重威胁。参与微生物腐蚀过程的包括真菌和藻类,以及各种类型的细菌。船舶船体、甲板、上层建筑、舱壁中大量使用的碳钢,常用作海水管系、热交换器、螺旋桨材料的铜合金以及在海洋平台中广泛应用的各种不锈钢,都不同程度地受到各类微生物的腐蚀而失效。因此,研究海洋微生物对金属材料的腐蚀机理,对提高船舶与海洋平台的寿命,增强服役安全性和可靠性具有重要意义。
1 海洋环境下的微生物腐蚀
1.1 微生物腐蚀概述
微生物腐蚀(Microbiological Induced Corrosion,MIC)是指金属或非金属表面因微生物的生命活动而受到的腐蚀破坏。海洋中的微生物接触到金属表面,分泌出胞外多糖(Exopolysaccharides)与金属表面粘附的有机物、无机物,甚至直接与金属基体发生反应,进一步形成胞外聚合物( Extracellular PolymericSubstances,EPS),细菌与EPS 一起附着于金属表面,形成生物膜。浸没在海水中的金属材料在数小时内就会覆盖一层生物膜,这层膜由各种细菌和微藻类以及它们的代谢产物组成,是发生微生物腐蚀的先决条件。生物膜内部的pH 值、溶解氧(Dissolved Oxygen,DO)、离子浓度与有机物含量等都与海水中有很大差异。金属材料与生物膜直接接触,微生物的代谢活动控制着金属表面电化学反应的类型与速率。
海洋中的微生物种类繁多,具有腐蚀能力的海洋细菌大多是铁、硫等元素循环过程的参与者,并改变金属表面的阴、阳极反应过程。根据细菌种类和代谢特点,可以将海洋中主要的腐蚀性细菌分为硫酸盐还原菌(Sulfate-reducing bacteria,SRB)、铁氧化细菌(Iron-oxidizing Bacteria,IOB)、铁还原细菌(IronreducingBacteria,IRB)、产酸菌(Acid-producingBacteria,APB)与产粘液菌(Slime-producing Bacteria,SPB)等,其中SRB 被认为是主要的腐蚀菌种,由它引起的金属材料腐蚀损失更是达到了微生物腐蚀损失的50%以上。
SRB 是在厌氧过程中占据主导地位的腐蚀菌种,有文献报道SRB 在非严格厌氧的环境下也可存活,所以SRB 也被认为是一种兼性厌氧菌[7]。最常见的SRB 包括脱硫弧菌属(Desulfovibrio sp.)与脱硫肠状菌属(Desulfotomaculum sp.),由于SRB 的作用,硫酸根离子被还原为S2-,进而在铁基金属表面留下典型的腐蚀产物FexS。SRB 是异养菌,可直接从金属基体获得电子以维持自身生命活动。IOB 能将亚铁离子(Fe2+)氧化为铁离子(Fe3+),并生成Fe(OH)3 沉淀,在金属表面产生氧浓差电池,引起局部腐蚀。IRB与IOB 相反,它将Fe3+还原为Fe2+,并进一步破坏金属表面的钝化膜以造成腐蚀[8]。SPB 通常是好氧的异养菌,常见的有假单胞菌(Pseudomonadaceae)与芽孢杆菌(Bacillaceae),其中Pseudomonadaceae 可以利用烃类作为能量来源,新陈代谢产生有机酸,并因此影响金属的腐蚀过程,生长旺盛时分泌大量的EPS并引起管路腐蚀、堵塞等问题。
1.2 船舶涉及的微生物腐蚀
严新平等将船舶摩擦分为外摩擦和内摩擦,MIC 与船舶的内、外摩擦均存在不同程度的协同作用。航行于海洋中的船体表面直接与海水接触,会有大量污损生物附着。杨宗澄等[11]调查了船舶表面生物污损情况,发现某些散货船的污损率可达到50%以上。多数船舶表面有防污漆保护,可以在一定程度上防止腐蚀。但大型污损生物,如藤壶等的长期附着,会破坏船壳表面的保护层,微生物迅速在船体表面破损处大量繁殖,使其表面产生脆弱的腐蚀产物层,受到海水冲刷时极易剥落,暴露出金属基体,从而加剧船体的外摩擦损失。Zarasvand 等从船体底部存在点蚀与黑色腐蚀产物的典型锈蚀部位取样分析,发现了大量SRB,其中多数为典型的腐蚀性菌属Desulfovibrio sp.。
与船舶外部环境相比,内部环境扰动小,各种舱室内温度范围在10~40 ℃之间,燃油、润滑油、舱内积水和生活污水的存在提供了丰富的营养物质,非常适合微生物生长。某些系统与舱室中,因含有油性污水、隔绝空气、清理困难等因素,形成缺氧且营养富集的环境,为一些厌氧菌的生长提供了条件,成为微生物腐蚀的多发部位。因此,船舶内部的微生物腐蚀相较于船舶外部更加普遍和严重,燃油系统、润滑系统、压载系统、液货舱以及各种液体管系都会发生不同程度的微生物腐蚀。
润滑油中的微生物能腐蚀舱壁,并随润滑油被输送至各摩擦副,加剧船舶主机各主要零部件、传动元件、支撑元件等的腐蚀磨损。船舶发电机中常用润滑脂作为润滑剂,其中的腐蚀微生物加速润滑脂变质失效,降低润滑性能与部件寿命。海洋微生物直接与水润滑尾轴承摩擦副相接触,微生物腐蚀会对轴承材料的摩擦学性能产生影响。船舶油舱底部常出现一层水,有时水层可占据油舱容积的20%,水中的微生物对舱底不断造成点蚀,形成局部缺陷,影响船舶的营运安全。有船舶曾发现碳钢板的点蚀速率达到了2 mm/a,考虑到舱室板厚度普遍介于10~15 mm之间,这样高的点蚀率造成了严重的安全隐患。有关调查显示,超过65%的船舶压载舱底部含有大量的沉积物,其中包含多种外来微生物,特别是厌氧菌与古生菌类(Archaea)可以在舱内缺氧环境下长期存活。陈德斌等报道了我国多艘舰艇舱底使用不到两年就发生了多处腐蚀穿孔,经检测后发现,舱底水中SRB数量是海水中的103~104 倍,蚀坑形貌多为阶梯型圆锥体,且腐蚀产物呈黑色泥状,有明显异味,认为SRB 是造成严重穿孔的主要原因。王毅等总结了近年来发生在船舶各部位中的微生物腐蚀(见表1),表明船舶上各种系统及结构部件均受到微生物腐蚀的影响。

1.3 海洋平台涉及的微生物腐蚀
海洋平台作为相对独立的系统在海洋环境下长期工作,根据其结构与功能可分为固定式、移动式与半固定式。海洋平台的腐蚀按纵向环境特点分为5 个区域,分别是海洋大气区、飞溅区、潮差区、全浸区与海泥区,微生物腐蚀主要存在于海洋大气区、全浸区以及海泥区。海洋大气区包括平台上的水、油、气管路以及动力系统和各种结构物,其环境与船舶内部环境相似,各种液体介质以及适宜的温度给微生物腐蚀创造了条件。全浸区的污损生物附着在桩腿上,微生物腐蚀使其界面的耐摩擦磨损性能大大降低。同时,微生物还能改变金属的腐蚀过程,引起局部腐蚀或穿孔,并在海流的冲刷作用下,导致腐蚀磨损加剧。海泥区由于深度大、海水中氧含量低,且海泥中存在H2S,厌氧菌特别是SRB 在腐蚀过程中占主导地位。
有关调查显示,在油气开采与运输储存过程中,输油管路、海水系统、造水系统等多处均会受到微生物腐蚀。Korenblum 等[23]研究了海洋采油平台海水注射系统中的微生物分布情况后发现,即使是经过灭菌处理的海水中依然存在包括SRB 在内的大量微生物,给海洋平台带来腐蚀加剧与石油酸化的威胁。一些近海港口或海岸由于污染造成了水质富营养化,Wiener 等[24]调查了污染水质中各物质的含量,发现有机碳、氮、氨以及有机硫化物的含量都很高,这种富营养化环境能促使微生物大量繁殖。
2 典型金属材料的微生物腐蚀
海洋运输船舶和海洋平台都是为海洋开发服务的关键装备。碳钢、不锈钢、铜合金是制造船舶和海洋平台的几种主要的金属材料。海洋微生物的代谢产物与生物膜会直接改变浸水材料表/界面的介质环境,将原有的海水/金属界面系统转变为海水/表面膜层/金属界面系统,加速金属材料的腐蚀。因此,碳钢、不锈钢、铜合金在苛刻海洋环境下的微生物腐蚀直接影响着船舶与海洋平台的可靠性与使用寿命。
2.1 碳钢
碳钢因较好的经济性与优良的焊接性、强度等综合性能,成为船舶上使用最广泛的材料,船体、各层甲板、舱壁、舱底板、上层建筑以及各种船舶结构物等普遍使用碳钢作为主要材料。一艘320 000 载重公吨位(DWT)的散货船碳钢用量可达到30 000 t 以上,100 000 DWT 级远洋船舶中有约90%的钢材是低碳钢。船舶常用的低碳钢(如Q235)及结构钢AH32、DH32、AH36 和DH36 等,强度高,但耐微生物腐蚀的能力较差。
海水中碳钢的腐蚀主要受到表面微环境中DO浓度、锈层厚度、EPS 理化性质、优势菌种更替、生物膜微观形态等因素控制。Melchers 等提出了一个模型,将碳钢在海水中的腐蚀过程分为需氧阶段(Mode1:oxygenated conditions dominant)与厌氧阶段(Mode2:anoxic conditions dominant),如图1 所示。在需氧阶段,碳钢的腐蚀速率取决于DO 的浓度高低,腐蚀产物累积并阻碍氧向碳钢表面扩散,同时在需氧菌的耗氧作用下形成缺氧环境,腐蚀开始进入厌氧阶段,微生物对碳钢的腐蚀主要发生在这一阶段。
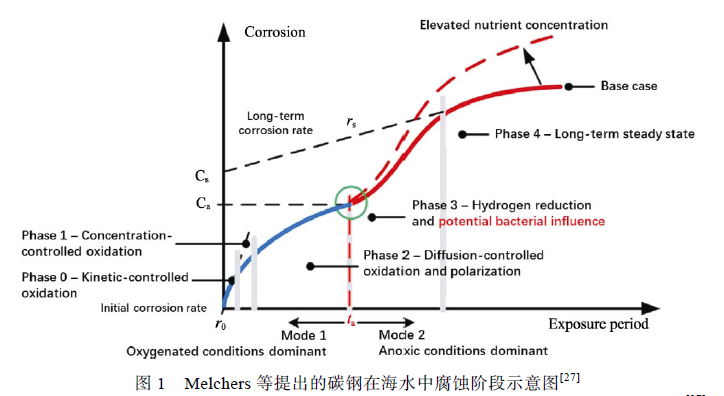
Lanneluc 等将碳钢试样在法国西部La Rochelle海港深度为1 m 的海水中浸泡8 周后发现,碳钢的腐蚀产物主要是铁的氧化产物( Fe3O4 、α-FeOOH、γ-FeOOH),同时在锈层内部发现了极少量的FeS,但是并没有发现SRB 存在于锈层中(可能是由于试验周期较短,导致SRB 的数量过少),说明腐蚀初期(1~8 周)主要发生的是由氧扩散主导的纯电化学腐蚀,但少量FeS 的出现表明厌氧菌已经开始影响腐蚀过程。根据Melchers 等提出的模型,此时碳钢表面的腐蚀主要处在需氧阶段,并开始向厌氧阶段过渡,但在需氧阶段并非没有微生物参与碳钢的腐蚀过程。Wu 等[29]研究了脱硫弧菌属(Desulfovibriosp.)与假交替单胞菌属(Pseudoalteromonas sp.)在天然海水中对低碳钢Q235 的腐蚀,发现DO 浓度是影响微生物腐蚀的重要因素。需氧菌Pseudoalteromonassp.耗尽了海水中的DO,当海水中仅存在Desulfovibrio sp.时,不会导致微生物腐蚀,这是由于高浓度DO 抑制了SRB 的生命活动。需氧菌Pseudoalteromonassp.的出现消耗了海水中的DO 并形成低氧环境,为Desulfovibrio sp.的生命活动提供了基本条件,引发微生物腐蚀。这说明在需氧阶段好氧菌阻碍DO 向碳钢表面扩散,起到抑制腐蚀的作用,同时DO 耗尽使锈层内优势菌种由需氧菌转变为厌氧菌,碳钢表面开始受到MIC 的影响。
微生物在不同生长阶段与生存环境中分泌的EPS 并非完全相同,EPS 理化性质差异影响着微生物对碳钢的腐蚀。董耀华等认为需钠弧菌(Vibrionatriegens)在DH32 表面分泌的EPS 与金属离子有络合作用,从而加剧碳钢的腐蚀。Dong 等提取了SRB 新陈代谢所产生的EPS,研究不同EPS 浓度对腐蚀的影响,发现低浓度的EPS 吸附在碳钢的表面,阻碍氧气与金属表面的接触,减缓了碳钢的腐蚀;高浓度的EPS 中含有大量氨基酸与核苷酸等负电基团,对二价铁离子(Fe2+)有明显的螯合作用,加速了碳钢表面的阳极溶解,使腐蚀更剧烈。Liu 等对比了不同生长时期IOB 的EPS 对低碳钢Q235 的腐蚀情况,随着IOB 的生长,EPS 的pH 值逐渐降低,腐蚀性增强,在碳钢表面EPS 与腐蚀产物形成的层状结构越来越疏松,碳钢所受腐蚀也更严重,这解释了为什么在Melchers 等提出的模型中,厌氧阶段碳钢腐蚀率高于需氧阶段。随着腐蚀产物不断累积,碳钢表面的离子浓度也会发生变化。Javed 等研究了铁离子浓度对SRB 附着与腐蚀的影响,在试验初期,铁离子浓度差异对碳钢表面的SRB 数量没有明显影响。随着试验进行至4~6 h,高铁离子浓度使得碳钢表面附着更多的SRB 且EPS 分泌量显着增多,这是由于脱硫弧菌细胞膜外层的脂多糖(lipopolysaccharide,LPS)与环境中的铁离子结合,促进了EPS 的分泌。
试验28 天后,碳钢腐蚀速率出现了巨大差异,高铁离子浓度试验组中,碳钢的腐蚀速率超过70 μm/a,而对照组(无铁离子)腐蚀速率仅有4 μm/a,两组相差17 倍之多。上述研究表明,高铁离子浓度能够显着促进SRB 的腐蚀,而碳钢的MIC 过程中能够产生大量的铁离子,即微生物腐蚀产物又进一步加剧了碳钢的MIC。碳钢为铁基金属材料,在含有SRB 的介质中会生成铁硫化物(FexS)与EPS 构成的复合生物膜。Xu 等认为碳钢表面的生物膜是非均匀的异相结构,其内层由多裂纹的腐蚀产物构成。Dong 等发现FexS 嵌入在疏松多孔的生物膜内,对腐蚀过程中的电子转移有良好的导通能力,使生物膜具备良好的电容特性。非均匀多孔异相生物膜使铁基金属表面存在浓度梯度,这种微环境差异造成表面各处金属阳极曲线发生变化,产生“自催化效应”,并造成更多的点蚀。同时,疏松生物膜中的微孔结构在碳钢MIC中起到关键作用,多孔生物膜下的碳钢表面电流分布并不均匀,这是由于腐蚀性离子通过生物膜上的微孔直接与碳钢基体接触所致。目前这种微孔结构的形成机制尚不清楚,有学者认为微生物分泌的酶导致了生物膜疏松多孔,在对EPS 进行酶失活处理后,生物膜失去了多孔特性,但有研究表明,含有生物酶的EPS 同样在碳钢表面形成了致密的生物膜,还有学者认为,酶能够利用葡萄糖产生H2O2,从而增大阴极电流密度。多种酶是微生物新陈代谢与金属腐蚀过程中不可或缺的活性物质,如氢化酶对MIC 过程中的氢消耗有决定性的作用,从生物酶的角度阐释腐蚀过程中的生物膜形态,为人们理解微生物腐蚀提供了一种新的思路。
实验室中针对单一菌种对碳钢的腐蚀已经进行了大量研究,但实际环境中微生物腐蚀通常是多种细菌联合作用的结果。近年来随着环保意识的提高,人们开始使用超低硫柴油(ultra low sulfur diesel,ULSD)作为船舶主机等大型动力装置的主要燃料。但超低硫柴油的使用伴随着更严重的局部腐蚀,船舶燃油柜、货油舱与油液管道等甚至出现因严重点蚀而失效的现象。Aktas 等[39]研究了碳钢在超低硫柴油使用环境下的微生物腐蚀,认为硫含量变化导致燃油对某些有降解能力的厌氧菌耐受程度降低,但这并没有显着影响碳钢的腐蚀。而ULSD 中存在的某些SRB如解烷烃脱硫簇杆菌(Desulfoglaeba alkanexedens),对其他类型SRB 的生长有促进作用,加剧了SRB 的新陈代谢,使碳钢受到更严重的局部腐蚀。菌种间具体的影响作用机制有待进一步研究。
2.2 不锈钢
不锈钢因良好的物理化学性能成为一种普遍采用的船舶与海洋工程材料。由于不锈钢具有优良的耐空泡腐蚀与耐冲蚀性能,美国、英国等在20 世纪80年代开始将不锈钢作为大型商船与高速舰船的螺旋桨材料,并取得了良好的效果。高速水翼艇的水翼要求材料强度高、韧性高与耐高速海水腐蚀性能好,双相不锈钢已成为水翼首选材料。特种运输船舶,如液化天然气运输船与腐蚀性化学试剂运输船中,不锈钢常用于制造货舱舱壁。舰船上各种接触海水的重要部件,如通海阀、波纹管与变速桨的机械连接部分等大量使用不锈钢材料。海洋油气开采平台在有H2S、硫酸气与氯化物的复杂海水环境中工作,其结构部件也大多采用不锈钢制造。
不锈钢表面在海洋中附着了生物膜后不再直接与海水接触,生物膜的微观形态、成膜过程变化都对材料的腐蚀有着很大的影响。Sheng 等[40]对比了不同SRB 菌种在316 不锈钢上的附着成膜情况后发现,脱硫脱硫弧菌(Desulfovibrio desulfuricans)在不锈钢表面形成的生物膜具有疏松网状的微观结构,不断地累积增厚,导致了更高的腐蚀速率。Cheng 等观察到Vibrio natriegens 在不锈钢表面的成膜过程与Desulfovibrio desulfuricans 有所不同,在成膜初期,因细菌生长繁殖,生物膜不断增厚,随后细菌数量下降,开始出现脱离现象(detachment phenomenon),同时形成具有疏松多孔结构的生物膜。这与碳钢表面的非均匀异相多孔生物膜很相似。微生物代谢产物也会加速不锈钢材料的腐蚀。Yuan 等对比了Desulfovibrio desulfuricans 代谢硫化物与无机硫化物对304 不锈钢的腐蚀情况发现,代谢硫化物对材料表面的腐蚀性更强,腐蚀产物、代谢硫化物与细菌共同组成了致密且粗糙的生物膜,代谢硫化物对不锈钢表面的氧化速度比无机硫化物更快。Chen 等对脱硫微杆菌(Desulfomicrobium sp.)的研究也得到了相似的结论,Desulfomicrobium sp.增大了不锈钢表面的粗糙程度与腐蚀速率,而无机硫化物在不锈钢表面形成的铁硫化物层更加均一稳定,腐蚀速率更小。
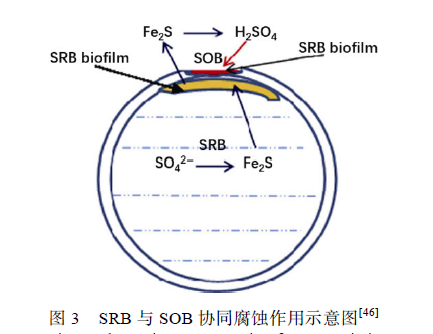
双相不锈钢(Duplex Stainless Steel,DSS)含有等量的奥氏体相(austenite(γ) phases)和铁素体相(ferrite (α) phases),同时具备铁素体不锈钢与奥氏体不锈钢的优点,并含有大量钝化元素,如铬(Cr)、钼(Mo)与镍(Ni),对Cl?腐蚀与硫化物腐蚀有着良好的抵抗能力。在过去几十年中,双相不锈钢,特别是2205 双相不锈钢(2205 DSS)被广泛应用在各种海洋工程结构物中,同时人们开始关注微生物对双相不锈钢造成的腐蚀。 Antony 等调查了Desulfovibrio desulfuricans对2205 DSS 的腐蚀情况发现,其表面同时受到了点蚀与缝隙腐蚀,缝隙腐蚀始于晶界处,并有选择性地向奥氏体相扩展,铁素体相所受腐蚀明显小于奥氏体相,在双相不锈钢表面形成了两种不同的微观形貌,说明奥氏体对SRB 腐蚀更加敏感,这可能与奥氏体中较低的Cr 与Mo 元素含量有关。在构件的某些部位如材料的焊接处,由于微生物的作用,耐蚀性优良的双相不锈钢也会发生异常严重的腐蚀。Liang 等报道了液态烃冷却器管路因腐蚀而失效的情况,在管内壁发现了较多点蚀,腐蚀产物中硫元素含量达到7.59%,说明海水中的SRB 直接参与了腐蚀过程。Liu等发现游艇上的2205 DSS 材料仅使用3 个月就在其焊接处出现了10 mm×60 mm 的异常蚀洞(如图2所示),提出导致这一异常腐蚀现象的原因是SRB 与硫氧化菌(SOB)的协同作用(如图3 所示)。海水中的厌氧菌SRB 在管道内壁形成生物膜并不断腐蚀2205 DSS 奥氏体相,同时将海水中的硫酸盐(SO4 2-)还原为Fe2S,蚀穿管壁后,海水中的需氧菌SOB 接触到空气,开始大量生长繁殖,SOB 又将SRB 的代谢产物Fe2S 氧化成有强烈腐蚀性的硫酸(H2SO4),在短时间内造成了异常严重的腐蚀现象。

微生物在不锈钢表面的成膜情况与金属材料的化学组分有关,Cr 与Mo 等元素使不锈钢表面在海水中形成钝化膜。不锈钢与碳钢虽同为铁基材料,但二者在MIC 中的表面层结构有显着差异。碳钢基体直接与疏松多孔的生物膜接触,相较于不锈钢来说,碳钢在微生物介质中的腐蚀速率要高得多。许多学者研究发现,不锈钢在发生微生物腐蚀后,形成了一种具有双层结构的表面膜,外层是由EPS 与腐蚀产物组成的生物膜,内层是钝化膜,说明不锈钢基体在MIC 过程中并不像碳钢那样直接与生物膜接触。
同时,微生物的生命活动能够改变不锈钢钝化膜的成分与结构,如SRB 的代谢产物H2S 能够破坏钝化膜,并产生铁硫化物。Dec 等对比了Desulfovibriodesulfuricans 在2205 DSS 与超级奥氏体不锈钢904L上的腐蚀情况发现,Desulfovibrio desulfuricans 在2205 DSS 表面的生物膜中含有大量EPS 与铁离子形成的牢固的有机金属键EPS—Fe,并加速不锈钢的阳极溶解。生物膜中含有大量的二硫阴离子S2 2-,对2205DSS 钝化膜有明显的硫化作用。不锈钢在海水中的抗蚀能力与Cr 元素含量密切相关,钝化膜内的Cr 含量同样受到微生物的影响。
Moradi 等研究了海洋菌假交替单胞菌(Pseudoalteromonassp.)在2205 DSS 上的腐蚀情况发现,生物膜中含有高浓度的Cl?,金属表面钝化膜内与蚀坑处表面Cr 含量降低,并随微生物生长,Cr 的流失量越大。但是Yuan 与Seyeux 等[53,54]的实验却得到了不同的结果,不锈钢受到微生物腐蚀后,其表面的铬含量反而增大,XPS 的结果显示,钝化膜内Cr 的主要化合物由Cr2O3 变为Cr(OH)3,认为Cr(OH)3 的抗蚀能力低于Cr2O3 是造成点蚀增多的主要原因。这些结果分别从不同的角度解释了不锈钢的MIC,前者认为微生物使Cr 流失,从而降低了不锈钢的抗蚀能力;后者则考虑了Cr 的化合物种类转变对MIC 的促进作用。这进一步反映出不锈钢相较于碳钢在微生物的影响下有更加复杂的腐蚀机制,一方面,钝化膜存在于生物膜与不锈钢基体之间,使表面层结构更加复杂,研究微生物对钝化膜的影响是理解不锈钢MIC 的关键;另一方面,钝化元素Cr 在MIC 过程中的含量变化一直存在争议。此外Cr 的抗微生物腐蚀能力尚未有研究证实。MIC 本质上是微生物在特定环境中的一种生存策略,深入研究Cr 元素在微生物活动中的转化过程,有助于从生物学的角度理解MIC 机理。
2.3 铜合金
早在19 世纪,铜合金就被应用于船舶与海洋工程领域,当时铜合金主要用作船体的保护层。在纯铜中加入镍元素可以显着提高材料的力学性能、耐蚀性与延展性,因此铜镍合金(又名白铜)在海洋工程领域备受青睐。在天然海水中,铜镍合金表面会迅速地生成主要由铜、镍的氧化物(Cu2O、Cu2(OH)3Cl、NiO、Ni(OH)2)构成的保护膜,这是铜镍合金具有较高耐蚀性的主要原因。海水淡化冷凝管、海水管道、舰船外壳等都大量使用铜镍合金作为主要材料。铜合金在海洋环境中具有良好的抗生物污损性能。Drach等选取了多种海洋工程领域常用的铜合金,在北大西洋海水中浸泡一年后发现,绝大多数试样表面没有出现宏观的生物污损,抗大型生物附着率高达100%。
铜离子通过破坏某些微生物细胞膜与DNA 蛋白质,从而具有一定的广谱抗菌作用。研究表明,在铁基合金中加入Cu 可以提高材料的抗菌性能,从而起到抑制微生物的腐蚀作用。Li 等研究了含铜2205双相不锈钢(2205-3%Cu DSS)在有海洋细菌绿脓杆菌(Pseudomonas aeruginosa)人造海水中的腐蚀情况,与2205 DSS 相比,2205-3%Cu DSS 上的生物膜数量显着降低,最大蚀坑深度也由4.83 μm 减小至1.53 μm。对铁基金属进行Cu 合金化功能改进后也发现,材料的生物膜贴附量与受点蚀程度明显降低。
因为铜在铁基合金中表现出良好的抗微生物腐蚀性能,那么以铜为主要基体元素的铜合金应该具备更加优良的抗菌与抗腐蚀能力,但大量的研究表明,铜合金在海洋微生物介质中的抗微生物贴附和抗蚀能力与含铜铁基合金有很大的差异。Krishnan 等对比了多种金属材料在海洋中微生物附着情况,发现铜合金表面的微生物数量虽少于不锈钢与碳钢,但铜离子的毒性并不能完全阻止微生物的生长,特别是革兰氏阳性菌(Gram-positive bacteria)的数量居多,如图4 所示,铜镍合金与黄铜表面的微生物附着量明显多于纯铜。

Javed 等报道了铜镍合金在SRB介质中的腐蚀仅次于碳钢,90Cu10Ni 在SRB 溶液中最大点蚀率达到82 μm/a,是在无菌溶液中均匀腐蚀速率(5 μm/a)的近17 倍, 而70Cu30Ni 的最大点蚀率也达到43 μm/a,这不仅远高于不锈钢在SRB 中的腐蚀速率(~0.1 μm/a),还出现了随着Cu 含量的增加,腐蚀反而加剧的现象。陈娟等观察SRB 对铜镍合金的腐蚀行为发现,在点蚀处Cu 的质量损失百分比仅是Ni 的1/4,在晶间腐蚀处Cu 的损失也远小于Ni 与Sn,这可能是由于EPS 对Ni 与Sn 有更强的络合作用,导致脱合金成分腐蚀,使铜镍合金表面出现大量点蚀与晶间腐蚀。Chen 等[62]研究了SRB 对金属铜的腐蚀发现,SRB 在Cu 表面能够快速生长繁殖,形成多相疏松的生物膜(如图5a 所示),在铜表面造成了明显的点蚀(如图5b 所示),分析腐蚀产物发现了大量的Cu2S。Sheng 等提出细菌EPS 与铜离子之间的络合反应是腐蚀过程中的关键,EPS 中羟基团的氧原子能够与环境中的游离态铜离子形成某种络合物,降低环境中的铜离子浓度,使细菌在一定程度上免受铜离子的毒性作用。上述研究结果解释了Krishnan 等发现的铜合金有较高微生物贴附量的现象,EPS 对Ni 与Sn 更加敏感,所以铜镍合金表面微生物附着量高于纯铜,而EPS 上的某些官能团,如羟基团中和了Cu 离子毒性,导致纯铜表面仍然有微生物贴附。

铜合金与不锈钢都属于钝化类金属材料,微生物在其表面的生命活动会改变钝化膜的物理化学状态。微生物分泌的EPS 从形态上可分为紧密结合型胞外聚合物(Tightly bound EPS,TB-EPS)与松散结合型胞外聚合物(Loosely bound EPS,LB-EPS),TB-EPS直接包裹微生物细胞形成类似胶囊结构,LB-EPS 则游离于EPS 的外层,且多呈黏液状。Bautista等提取了海洋假单胞菌(Pseudomonas)分泌的LB-EPS 并贴附在铜镍合金表面,发现钝化膜厚度明显降低,认为LB-EPS 能阻碍Cl-与Cu 接触,从而具有抑制腐蚀的效果。但仅从EPS 的角度去理解铜合金的MIC 是不全面的,因为在含有微生物的介质中,生物膜状态远比均匀贴附要复杂得多。Fu 等发现,SRB 新陈代谢所产生的硫化物具有强烈的腐蚀性,能使钝化膜持续溶解,黑色腐蚀产物与EPS 以及微生物细胞共同组成具有疏松多孔特性的生物膜,这与碳钢表面生物膜的不均匀异相结构非常相似。Tuba等认为,由于细菌对毒性离子(Cu2+)的天然反应,EPS 的含量增多,并与大量细菌以及腐蚀产物聚集在一起,所形成的生物膜面积变小但更厚,可能有助于微生物抵抗铜离子毒性。Huang 等观察到铜合金表面的生物膜分布呈现出非连续的团聚现象,明显区别于碳钢与不锈钢表面生物膜的连续分布,这证实了Tuba 与Sheng 对微生物抵抗Cu 毒性机制的猜测。虽然微生物分泌的EPS 能与Cu 发生某种络合反应,但并未形成类似于铁基金属表面上的牢固有机金属键(EPS—Fe),所以铜合金表面的生物膜更加疏松脆弱,导致钝化膜被完全破坏,直至暴露基体金属,造成局部腐蚀。表2 中对比了铜合金、碳钢与不锈钢在微生物介质中的腐蚀特征,铜合金钝化膜在SRB 介质中受到很大程度的破坏,其中的氧化铜完全转变为Cu2S,使钝化膜失去对基体的保护能力,这也就解释了为什么铜合金受到的微生物腐蚀远比不锈钢严重。

目前人们对Cu 的抗菌机理并无统一定论,仅提出了几种可能的机制:1)铜离子直接接触杀菌;2)催化作用杀菌;3)破坏细菌酶系统杀菌;4)凝固细菌蛋白质杀菌。从上述研究工作来看,微生物在对铜合金的腐蚀过程中,对Cu 的毒性有一定的抵御策略,Cu 不仅未起到明显的抗菌效果,铜合金在海水中的抗腐蚀能力也因微生物附着而急剧降低[47,58-69]。对于铜合金与含铜铁基合金在抗微生物腐蚀方面的较大差异值得深入研究,这不仅为Cu 的抗菌机制研究提供了一个新的视角,同时对如何增强材料的抗微生物腐蚀性能具有重要的指导意义。
3 微生物腐蚀机理研究进展
金属材料的电化学腐蚀经过多年的研究,已经有比较透彻的机理解释,微生物腐蚀的机理也吸引了众多的学者去开展这方面的研究工作,阴极去极化理论和微生物电化学腐蚀理论是目前最主要的用于微生物腐蚀机理解释的理论。
3.1 阴极去极化理论
阴极去极化理论是早期学者通过研究SRB 对碳钢的腐蚀所提出的[71,72]。这个理论认为,微生物能通过消耗金属表面在水中形成的氢,并降低局部分压,起到去极化剂的作用,加速金属阳极溶解,同时电化学意义上的阴极氢也能促进微生物的生长。SRB 附着在碳钢表面后捕获阴极上的吸附氢(Hads),在硫酸根离子(SO4 2 -)的还原过程中,Hads 被消耗(见式(1)),生成S2-,氢膜被破坏后无法阻止Fe2+由金属基体向生物膜扩散,Fe2+与S2-生成FeS(见式(2)),沉淀在生物膜中,形成疏松多孔的表面层。
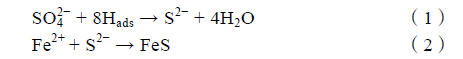
用阴极去极化理论解释SRB 对碳钢的腐蚀机理得到了众多学者的支持,在碳钢表面观察到的不均匀异相生物膜以及对腐蚀产物的分析,都证实了阴极去极化理论的可行性。SRB 分泌的氢化酶在消耗氢的过程中起到关键的作用。Liu 等将生物膜进行处理,使酶失去活性,发现碳钢表面的层状结构变得紧密,腐蚀产物中的铁硫化物含量显着降低,说明氢化酶控制着S2-的生成。
然而仅用阴极去极化理论无法解释所有的微生物腐蚀现象。按照阴极去极化理论,SRB 在没有碳源作为电子供体的情况下,无法将海水中的SO4 2?还原为S2-,对碳钢的腐蚀应该有所减轻,但Xu 等发现,SRB 在有机碳源匮乏的情况下,对碳钢的腐蚀反而更加剧烈。当环境介质中缺乏90%的有机碳源时,脱硫弧菌(Desulfovibrio vulgaris)对碳钢的平均点蚀深度达到7 μm,是碳源充足时的近两倍。Chen 等进一步研究了SRB 在有机电子供体缺乏的条件下对碳钢的腐蚀情况,随着溶液中营养物质的不断消耗,游离态细菌比例从实验初期的(92±5)%逐渐下降至(6±4)%,游离态的SRB 转而吸附在碳钢表面,局部腐蚀也更加严重。另外,有研究表明,微生物活动能够改变不锈钢钝化膜内的化合物成分与元素含量,并阻碍不锈钢表面的保护层再生,因此不锈钢的MIC机理更加复杂。
3.2 微生物电化学腐蚀理论
由于阴极去极化理论的局限性,学者们试图从其他角度去理解MIC。有研究指出,微生物燃料电池(Microbial fuel cells,MFCs)中存在细胞外电子传递机制,微生物细胞可以通过生物纳米导线、细胞膜功能蛋白、活性电子中介体等多种方式,将金属基体元素的电子转移至细胞内,作为生命活动的能量来源。
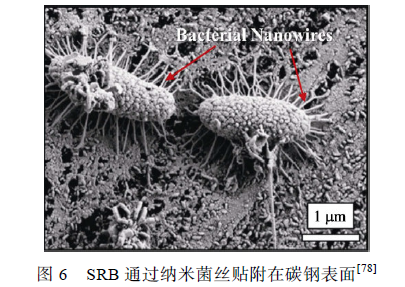
SRB 细菌胞体能分泌出多条与金属表面紧密结合的纳米菌丝(Bacterial nanowires),使细菌牢固地贴附在碳钢表面,如图6 所示。Yu 等证明了SRB 的电活性,认为SRB 可以从极化0.61 V(vs. SHE)的石墨电极中直接或间接地获得电子。Guan 等发现0.85 V(vs. SCE)的极化电位更有利于SRB 生长,提出SRB 可以从此电位下的电极中获得电子,促进金属腐蚀。Xu 等提出了生物阴极催化硫酸盐还原机理,认为SRB 可以通过纳米菌丝或某些特异蛋白质直接利用金属基体元素作为电子供体,这个过程就表现为微生物对金属产生腐蚀作用。根据这个理论,微生物优先利用环境介质中的营养物质作为电子供体,随着腐蚀产物和生物膜的累积增厚,环境中的有机碳源难以被生物膜底层的微生物捕获,这时金属基体元素如Fe 就成为唯一的电子供体。阳极反应为Fe失去电子被氧化为Fe2+(见式(3)),阴极反应为SO4 2-被还原为S2-(见式(4)),与阴极去极化理论中的阴极反应相似,不同的是生物阴极催化硫酸盐还原理论认为阴极过程直接发生在微生物细胞内,传统观点中的物理阴极被发展为生物阴极。
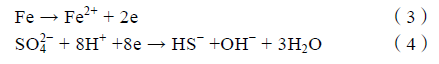
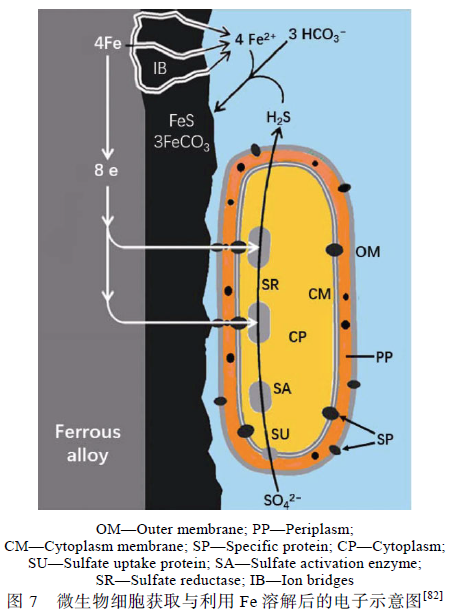
Venzlaff 与Enning 等[82,83]强调微生物电化学过程对金属腐蚀的影响,提出了与阴极催化硫酸盐还原机理相似的电化学微生物腐蚀理论,并给出了电子与各离子在微生物细胞内的转移过程,见图7。细胞外膜(OM)、外周胞质(PP)与细胞质膜(CM)上均存在特异蛋白质(SP),能够将细菌胞外的电子传递至细胞质(CP)内,同时CM 上的硫酸盐摄取蛋白质(SU)捕获环境介质中的SO4 2 -,通过细胞质内的硫酸盐活化酶(SA)与硫酸盐还原酶(SR)利用金属失去的电子将SO4 2-还原为H2S,Fe2+能通过生物膜上的离子桥(IB)与H2S 接触生成腐蚀产物FeS。他们提出的离子桥(IB)结构与文献[34]中报道的碳钢表面生物膜微孔类似。Yu 等证实了SRB 的电子传递在碳钢的腐蚀中起到重要作用,并为SRB 与碳钢之间的电子传递提供了依据。Xu 与Chen 等的研究结果以及生物膜具有良好导电性的发现[35]也为微生物电化学腐蚀理论提供了有力证据。
阴极去极化理论可以被认为是微生物电化学腐蚀理论的一种特殊情况。当微生物能够分泌氢化酶时,将利用金属表面的H 作为电子供体,并起到阴极去极化的作用,但对于无法分泌氢化酶或无法直接利用环境中碳源的微生物来说,可以通过多种方式从金属基体获取电子,以支持生命活动。微生物电化学腐蚀理论将电化学与微生物能量学联系在一起,具有开创性,使人们对MIC 有了全新的认识。但这个理论同样存在缺陷,比如Cu 的SRB 腐蚀就很难用微生物电化学的机制去解释。根据腐蚀热力学,腐蚀发生的前提条件是氧化-还原反应的电化学亲和势A 为大于零的正值(见式(5))。

式中:A 为电化学亲和势;n 为电子化学计量系数;F 为法拉第常数,约为96 494 C;E02与E01 分别为阴极与阳极的平衡电位。
Cu 被分别氧化为Cu+与Cu2+的阳极反应中,相对饱和甘汞电极(vs. SHE)的标准电位分别为522 mV与345 mV,若将Cu 的两种氧化反应作为SO4 2 -还原过程的阳极计算,得到了小于零的电化学亲和势,这与腐蚀热力学的观点相悖,说明微生物可能无法直接利用Cu 作为电子供体。Fu 等认为铜晶体(Cu(crystal))虽然无法参与微生物细胞内的阴极过程,但可以与新陈代谢产物S2-反应,生成硫化亚铜沉淀(Cu2S(crystal))(见式(6)),H 作为电子转移载体出现,且最终生成氢气(H2(g))。这说明Cu 在SRB 作用下的腐蚀过程,既不完全符合阴极去极化理论中消耗氢的观点,又无法用微生物的电化学腐蚀来解释。
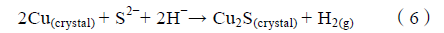
从目前的研究来看,对Cu 的MIC 机理倾向于用微生物代谢产物具有较强腐蚀性来解释,如SRB 产生的H2S,APB 与SPB 分泌的有机酸等。微生物腐蚀是一个复杂的过程,无论是阴极去极化理论或是近年来提出微生物电化学腐蚀理论,都不会在MIC 中单独起作用,如硫还原地杆菌细胞外膜上有丝状的菌毛蛋白(Geopili),能在细胞与金属之间传递电子,脱氮硫杆菌无法直接利用金属被氧化后的失去电子,但可以通过某种导电的纳米颗粒将硫还原地杆菌获取的电子转移至细胞内以供生命活动,对金属造成间接性的腐蚀。除了菌种间的协同与竞争作用,生物膜的附着状态、微观结构、膜内沉积的腐蚀产物以及表面微环境的差异,都能影响微生物对金属的腐蚀,生存环境的改变也能导致同种微生物产生不同的腐蚀效果。
针对碳钢、不锈钢和铜合金这几种最主要的海洋金属材料,上述两种理论都在一定程度上可以对他们的微生物腐蚀机理做出一些解释。碳钢、不锈钢与铜合金的MIC 必定伴随着金属基体的溶解,根据上述理论,微生物主要通过以下几种方式影响溶解过程:
1)改变金属阳极失去电子的转移过程与电子受体,微生物电化学腐蚀理论就是从这个角度阐释了碳钢的MIC 过程;2)破坏金属表面原有的保护膜,碳钢、不锈钢与铜合金的MIC 中都体现出了微生物对保护膜的影响,阴极去极化理论认为SRB 消耗了碳钢表面的氢膜,不锈钢与铜合金的钝化膜也受到了不同程度的破坏;3)腐蚀性新陈代谢产物加速金属溶解,铜合金基体在SRB 海水中的局部腐蚀就主要受代谢产物H2S 的控制。然而,上述两种理论还不能完整地对海洋材料的微生物腐蚀机理做出完全令人信服的解释,这也充分说明了微生物腐蚀过程的复杂性,还亟需进一步开展这方面的研究,来继续完善微生物腐蚀机理。
4 展望
海洋环境中金属材料的微生物腐蚀研究越来越受到人们的关注,大量学者在微观层面对微生物腐蚀机理进行了研究,提出了一些理论模型并试图解释金属材料发生微生物腐蚀的过程,并取得了一定的研究成果。但海洋微生物腐蚀过程极其复杂,它不是一个简单的电化学过程,其研究涉及到海洋科学、微生物学、界面化学、金属材料等多个学科领域的交叉,而且目前的实验技术手段还不足以完全掌握微生物腐蚀机理,加之复杂的海洋工况环境,仍有诸多问题值得研究与探讨:
1)不同菌种间的协同作用机理研究。海洋环境中微生物种类繁多,对单一菌种的实验研究难以反映出金属材料在实际工况下的腐蚀,菌种间的相互作用机理、协同作用如何影响腐蚀过程,与单一菌种的腐蚀有何区别,这些将成为今后重要的研究方向。
2)非铁基金属的MIC 机理研究。针对铁基金属的MIC 已进行了大量研究,得到了具有代表性的几种腐蚀理论,但这些理论无法完全解释非铁基金属,如铜合金的MIC 腐蚀机理还没有一个完整的机制理论,对此进行深入研究能进一步理解MIC 过程。
3)微生物对海洋耐蚀材料的影响。海洋耐蚀合金相较于一般的金属材料具有更复杂的化学成分与金相结构,对海水腐蚀有良好的抵抗能力,但对微生物的耐蚀性如何,耐蚀元素如Cr 是如何影响微生物的成膜与腐蚀过程等,都有待进一步研究。
4)微生物腐蚀与摩擦磨损的协同作用。目前多数有关海洋环境下摩擦副摩擦磨损行为的研究未考虑微生物腐蚀的影响,两者间的相互促进或抑制作用机理的研究对提高材料在海洋中的使用性能具有重要指导意义。
5)微生物与宏观污损生物的协同作用。宏观污损生物附着于金属表面,加速材料的局部腐蚀,其下的锈层中往往存在微生物群落,宏观污损生物与微生物的共存如何影响腐蚀,两者间的协同机理尚不明确。